Los elementos internos son:
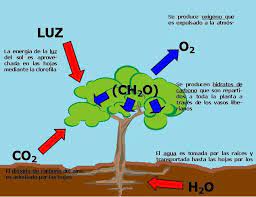

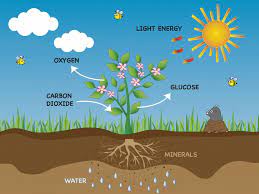
La fotosíntesis es el proceso de obtención de energía de plantas, algas y ciertas bacterias por medio de la luz solar. También recibe el nombre de función clorofílica. La fotosíntesis o función clorofílica es un proceso químico que consiste en la conversión de materia inorgánica a materia orgánica gracias a la energía que aporta la luz solar. En este proceso, la energía lumínica se transforma en energía química estable, siendo el NADPH (nicotín adenín dinucleótido fosfato) y el ATP (adenosín trifosfato) las primeras moléculas en la que queda almacenada esta energía química.
Con posterioridad, el poder reductor del NADPH y el potencial energético del grupo fosfato del ATP se usan para la síntesis de hidratos de carbono a partir de la reducción del dióxido de carbono (CO2). La vida en nuestro planeta se mantiene fundamentalmente gracias a la fotosíntesis que realizan en el medio acuático las algas, las cianobacterias, las bacterias rojas, las bacterias púrpuras, bacterias verdes del azufre, y en el medio terrestre las plantas, que tienen la capacidad de sintetizar materia orgánica (imprescindible para la constitución de los seres vivos) partiendo de la luz y la materia inorgánica. De hecho, cada año los organismos fotosintetizadores fijan en forma de materia orgánica en torno a 100 000 millones de toneladas de carbon
Técnicamente, el proceso de la fotosíntesis convierte el dióxido de carbono (CO2) y el agua (H2O) en glucosa (C6H12O6), es decir, convierte sustancias inorgánicas en sustancias orgánicas Los organismos que poseen la capacidad de la fotosíntesis se llaman autótrofos en tanto que no dependen de otros seres vivos para alimentarse. Dentro de esta categoría, clasifican concretamente como fotoautótrofos, pues obtienen energía a partir de la luz solar.
La palabra fotosíntesis es de origen griego photo que significa “luz”, syn que es equivalente a “con” y thesis que expresa “conclusión o posición”.
Los organismos que tienen la capacidad de llevar a cabo la fotosíntesis son llamados, fotoautótrofos (otra nomenclatura posible es la de autótrofos, pero se debe tener en cuenta que bajo esta denominación también se engloban aquellas bacterias que realizan la quimiosíntesis) y fijan el CO2 atmosférico. En la actualidad se diferencian dos tipos de procesos fotosintéticos, que son la fotosíntesis oxigénica y la fotosíntesis anoxigénica. La primera de las modalidades es la propia de las plantas superiores, las algas y las cianobacterias, donde el dador de electrones es el agua y, como consecuencia, se desprende oxígeno. Mientras que la segunda, también conocida con el nombre de fotosíntesis bacteriana, la realizan las bacterias purpúreas y verdes del azufre, en las que el dador de electrones es el sulfuro de hidrógeno (H2S), y consecuentemente, el elemento químico liberado no será oxígeno sino azufre, que puede ser acumulado en el interior de la bacteria, o en su defecto, expulsado al agua.
Los elementos internos son:
En la Antigua Grecia, el filósofo Aristóteles propuso una hipótesis que sugería que la luz solar estaba directamente relacionada con el desarrollo del color verde de las hojas de las plantas, pero esta idea no trascendió en su época, quedando relegada a un segundo plano.
A su vez, la idea de que las hojas de las plantas asimilaban el aire fue propuesta por Empédocles, y descartada por Aristóteles y su discípulo Teofrasto, quien sostenía que todo el «alimento» de las plantas provenía de la tierra. De hecho, esas ideas no volvieron a ser recuperadas hasta el siglo XVII, cuando el considerado padre de la fisiología vegetal, Stephen Hales, hizo mención a las citadas hipótesis, y afirmó que el aire que penetraba por las hojas en las plantas era empleado por ellas como fuente de alimento.
Durante el siglo XVIII comenzaron a surgir trabajos que relacionaban los incipientes conocimientos de la química con los de la biología. En la década de 1770, el clérigo inglés Joseph Priestley (a quien se le atribuye el descubrimiento del O2) estableció la producción de oxígeno por los vegetales, reconociendo que el proceso era, de forma aparente, el inverso de la respiración animal, que consumía tal elemento químico. Fue Priestley quien acuñó la expresión de aire deflogisticado para referirse a aquel que contiene oxígeno y que proviene de los procesos vegetales, así como también fue él quien descubrió la emisión de dióxido de carbono por parte de las plantas durante los periodos de penumbra, aunque en ningún momento logró interpretar estos resultados.
En 1778, el médico neerlandés Jan Ingenhousz dirigió numerosos experimentos dedicados al estudio de la producción de oxígeno por las plantas (muchas veces ayudándose de un eudiómetro), mientras se encontraba de vacaciones en Inglaterra, para publicar al año siguiente todos aquellos hallazgos que había realizado durante el transcurso de su investigación en el libro titulado Experiments upon Vegetables. Algunos de sus mayores logros fueron el descubrimiento de que las plantas, al igual que sucedía con los animales, viciaban el aire tanto en la luz como en la oscuridad; que cuando los vegetales eran iluminados con luz solar, la liberación de aire cargado con oxígeno excedía al que se consumía y la demostración que manifestaba que para que se produjese el desprendimiento fotosintético de oxígeno se requería de luz solar.
También concluyó que la fotosíntesis no podía ser llevada a cabo en cualquier parte de la planta, como en las raíces o en las flores, sino que únicamente se realizaba en las partes verdes de esta. Como médico que era, Jan Ingenhousz aplicó sus nuevos conocimientos al campo de la medicina y del bienestar humano, por lo que también recomendó sacar a las plantas de las casas durante la noche para prevenir posibles intoxicaciones.
De todas las células eucariotas, únicamente las fotosintéticas presentan cloroplastos, unos orgánulos que usan la energía de la luz para impulsar la formación de ATP y NADPH, compuestos utilizados con posterioridad para el ensamblaje de azúcares y otros compuestos orgánicos. Al igual que las mitocondrias, cuentan con su propio ADN y se han originado a partir de bacterias simbióticas intracelulares (teoría endosimbiótica).
Los cloroplastos se distinguen por ser unas estructuras polimorfas de color verde, siendo la coloración que presentan consecuencia directa de la presencia del pigmento clorofila en su interior. Los cloroplastos están delimitados por una envoltura formada, en la mayoría de las algas y en todas las plantas, por dos membranas (externa e interna) llamadas envueltas, que son ricas en galactolípidos y sulfolípidos, pobres en fosfolípidos, contienen carotenoides y carecen de clorofila y colesterol. En algunas algas, las envueltas están formadas por tres o cuatro membranas, lo que se considera prueba de que se han originado por procesos de endosimbiosis secundaria o terciaria. Las envueltas de los cloroplastos regulan el tráfico de sustancias entre el citosol y el interior de estos orgánulos, son el lugar de biosíntesis de ácidos grasos, galactolípidos y sulfolípidos y son el lugar de reconocimiento y que contiene los elementos necesarios para permitir el transporte al interior de los orgánulos de las proteínas de cloroplastos codificadas en el núcleo celular.
En las plantas superiores, la forma que con mayor frecuencia presentan los cloroplastos es la de disco lenticular, aunque también existen algunos de aspecto ovoide o esférico. Con respecto a su número, se puede decir que en torno a cuarenta y cincuenta cloroplastos coexisten, de media, en una célula de una hoja; y existen unos 500.000 cloroplastos por milímetro cuadrado de superficie foliar. No sucede lo mismo entre las algas, pues los cloroplastos de estas no se encuentran tan determinados ni en número ni en forma. Por ejemplo, en el alga Spirogyra únicamente existen dos cloroplastos con forma de cinta en espiral, y en el alga Chlamydomonas, solamente hay uno, de grandes dimensiones.
En el interior y delimitado por la membrana plastidial interna, se ubica una cámara que alberga un medio interno con un elevado número de componentes (ADN plastidial, circular y de doble hélice, plastorribosomas, enzimas e inclusiones de granos de almidón y las inclusiones lipídicas); es lo que se conoce por el nombre de estroma. Inmerso en él se encuentran una gran cantidad de sáculos denominados tilacoides, cuya cavidad interior se llama lumen o espacio tilacoidal. En las membranas de los tilacoides se ubican los complejos proteínicos y complejos pigmento/proteína encargados de captar la energía lumínica, llevar a cabo el transporte de electrones y sintetizar ATP.
Los tilacoides pueden encontrarse como vesículas alargadas repartidos por todo el estroma (tilacoides del estroma), o bien, pueden tener forma discoidal y encontrarse apilados originando unos montones, denominados grana (tilacoides de grana).
La más importante función realizada en los cloroplastos es la fotosíntesis, proceso en la que la materia inorgánica es transformada en materia orgánica (fase oscura) empleando la energía bioquímica (ATP) obtenida por medio de la energía solar, a través de los pigmentos fotosintéticos y la cadena transportadora de electrones de los tilacoides (fase luminosa). Otras vías metabólicas de vital importancia que se realizan en el estroma, son la biosíntesis de proteínas y la replicación del ADN.
La energía lumínica que absorbe la clorofila excita a los electrones externos de la molécula, los cuales pueden pasar a otra molécula adyacente (separación de cargas). Es decir, la energía lumínica impacta en los pigmentos fotosintéticos presentes en las membranas tilacoidales.
Cuando la luz es absorbida por uno de estos pigmentos de la antena, la energía se traspasa de una molécula a otra del fotosistema mediante resonancia inducida hasta que alcanza una forma especial de “clorofila a” que forma parte del “centro reactivo" del fotosistema. Se produce una corriente electrónica (transporte de electrones) en el interior del cloroplasto a través de la cadena de transporte de electrones. El fotosistema II absorbe luz con longitudes de onda de hasta 680 nm y el fotosistema I absorbe luz con longitudes de onda de hasta 700 nm. La energía (procedente de la luz) de los electrones que se transportan es empleada indirectamente en la síntesis de ATP mediante la fotofosforilación (precisa transporte de protones desde el lumen tilacoidal al estroma), y directamente en la síntesis de NADPH (el NADP recibe los electrones procedentes del agua, al final de la cadena de transporte y se reduce a NADPH). Ambos compuestos son necesarios para la siguiente fase o Ciclo de Calvin, donde se sintetizarán los primeros azúcares que servirán para la producción de sacarosa y almidón. Los electrones que ceden las clorofilas son repuestos mediante la oxidación del H2O, proceso en el cual se genera el O2 que las plantas liberan a la atmósfera.
Existen dos variantes de fotofosforilación: acíclica y cíclica, según el tránsito que sigan los electrones a través de los fotosistemas. Las consecuencias de seguir un tipo u otro estriban principalmente en la producción o no de NADPH y en la liberación o no de O2.
El proceso de la fase luminosa, supuesto para dos electrones, es el siguiente: Los fotones inciden sobre el fotosistema II, excitando y liberando dos electrones, que pasan al primer aceptor de electrones, la feofitina. Los electrones los repone el dador último de electrones, el dador Z, con los electrones procedentes de la fotólisis del agua en el interior del tilacoide (la molécula de agua se divide en 2H+ + 2e- + 1/2O2)Los protones de la fotólisis se acumulan en el interior del tilacoide, y el oxígeno es liberado.

Los electrones pasan a una cadena de transporte de electrones, que invertirá su energía liberada en la síntesis de ATP. ¿Cómo? La teoría quimioosmótica nos lo explica de la siguiente manera: los electrones son cedidos a las plastoquinonas, las cuales captan también dos protones del estroma. Los electrones y los protones pasan al complejo de citocromos bf, que bombea los protones al interior del tilacoide. Se consigue así una gran concentración de protones en el tilacoide (entre estos y los resultantes de la fotólisis del agua), que se compensa regresando al estroma a través de las proteínas ATP-sintasas, que invierten la energía del paso de los protones en sintetizar ATP. La síntesis de ATP en la fase fotoquímica se denomina fotofosforilación.
Los electrones de los citocromos pasan a la plastocianina, que los cede a su vez al fotosistema I. Con la energía de la luz, los electrones son de nuevo liberados y captados por el aceptor A0. De ahí pasan a través de una serie de filoquinonas hasta llegar a la ferredoxina. Esta molécula los cede a la enzima NADP+-reductasa, que capta también dos protones del estroma. Con los dos protones y los dos electrones, reduce un NADP+ en NADPH + H+.
El balance final es: por cada molécula de agua (y por cada cuatro fotones) se forman media molécula de oxígeno, 1,3 moléculas de ATP, y un NADPH + H+.